我猜你枪里没子弹什么意思啊(你猜我枪里有没有子弹表情包)
导语:你猜我枪里有多少子弹——囊泡回收
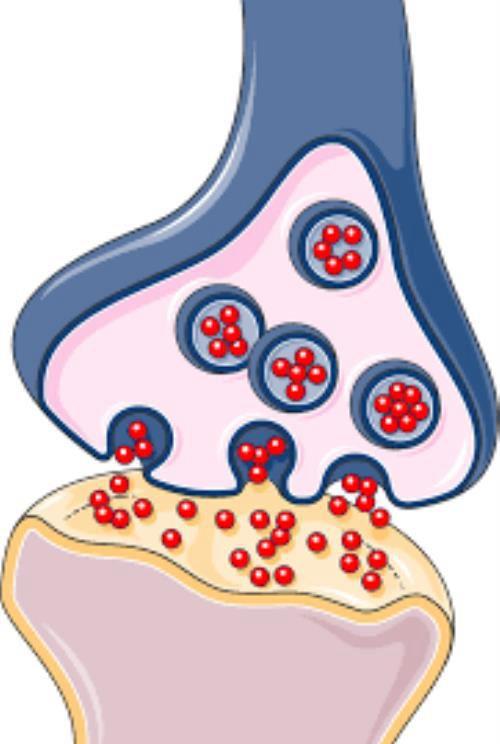
当突触前神经元像机关枪一样“突突突”地发射名叫“神经递质”的子弹时,这把机关枪早已为自己找好了新的子弹。
囊泡回收神经元之间的交流需要突触前神经元释放神经递质,这是一种包裹在囊泡里的化学物质。神经递质释放的瞬间,正像子弹脱出弹夹,射出枪膛。随子弹出壳,这些装满弹药的弹夹——囊泡,也随之消逝在一望无际的细胞膜里。
当神经递质释放时,囊泡与细胞膜融合,细胞膜面积变大,细胞内的囊泡数量必然会减少。
那么,神经元是如何保证自己有取之不尽、用之不竭的囊泡的呢?
原来,像机关枪一样的神经元之所以能够肆无忌惮地开枪——发放动作电位,是因为它早已想好了补充新子弹的方法。这个方法就是——囊泡回收。
细胞会通过胞吞的方式,将失去的囊泡从细胞膜上夺回来。也就是说,细胞膜会向内部凹陷,形成新的囊泡。这个过程就是囊泡回收。
网格蛋白介导的内吞(Clathrin-Mediated Endocytosis,CME)经典的囊泡回收方式是由网格蛋白(Clathrin)介导的,因此也称为网格蛋白介导的内吞(Clathrin-Mediated Endocytosis,CME)。
简单的讲,细胞膜上的一片膜会向内凹陷,由网格蛋白(Clathrin)包裹形成囊泡雏形,通过动力蛋白(Dynamin)切断该凹陷与细胞膜之间的连接处,使囊泡回收到细胞内。在这个过程中,网格蛋白像一件外衣一样,装配起来,包裹在新形成的囊泡上。待囊泡与细胞膜脱离后,需要解装配去掉网格蛋白的外衣。具体过程参见:着衣又去衣,这是胞吞的仪式。
尽管网格蛋白介导的内吞(Clathrin-Mediated Endocytosis,CME)是囊泡回收的一种主要方式,然而,由于这种方式需要网格蛋白的装配与解装配,因此耗时很长,约30秒才能完成整个囊泡的内吞过程。
在静息状态下,细胞自发的发放动作电位和释放神经递质。没有外部压力的神经元无需迅速回收囊泡,只需用网格蛋白介导的内吞作为默认的途径回收囊泡即可。
网格蛋白介导的内吞就像重新制作新子弹,运到战场上,将子弹装进机关枪的弹夹。这种回收囊泡的方式就像是战场的大后方,在不断给神经元这个机关枪供应子弹和弹夹。
然而,当细胞接受刺激或面对压力时,会以极高的速率排出囊泡里的神经递质。此时,这些刺激就像扣动了机关枪的扳机,使得囊泡里装的“子弹”被快速释放出去。在这种情况下,打完“子弹”的细胞需要快速找到新的子弹,以应对接下来的挑战。
因此,只有一种网格蛋白介导的内吞来回收囊泡,显然还远远不够。细胞需要多种内吞方式来满足快速补充囊泡的目的。
囊泡回收的方式除网格蛋白介导的内吞外,细胞还可以通过多种方式,例如,大胞饮(macropinocytosis)、吻了就跑(kiss-and-run)、大泡内吞(Activity Dependent Bulk Endocytosis,ADBE)、吞蛋白介导的快速内吞(Fast Endophilin-Mediated Endocytosis, FEME),超快速内吞(Ultrafast endocytosis)等进行囊泡回收。
在这几种方式中,囊泡回收的速度是很不一样的。例如,网格蛋白介导的内吞速度约30秒,“吻了就跑”速度约1-2秒,大泡内吞速度约10秒到1分钟,吞蛋白介导的快速内吞速度约为3秒,超快速内吞速度约为470毫秒。
下面,我们主要介绍其中速度适中的一种囊泡回收途径——吞蛋白介导的快速内吞。
吞蛋白介导的快速内吞吞蛋白介导的快速内吞主要在非神经元细胞中某些受体被激活时启动。
例如,当神经内分泌细胞受到配体刺激时,激活肾上腺素能受体,诱发囊泡释放。其中,β1肾上腺素能受体在被激活后,能够引发吞蛋白介导的快速内吞。相反,β2肾上腺素能受体则不能引发该途径。
有了受体的激活后,相比网格蛋白这个运输子弹的大后方,吞蛋白介导的快速内吞(Fast Endophilin-Mediated Endocytosis, FEME)就像有个助手专门在给弹夹装子弹,速度快了10倍。
网格蛋白介导的内吞像个乌龟,需要30秒甚至几分钟的时间,才能完成囊泡回收的全部过程。然而,吞蛋白介导的快速内吞像是兔子,只需要3秒,就能完成“吞”的动作。
两种方式相比就像龟兔赛跑。乌龟缓缓道:开始了吗?只见兔子早已结束战斗,站在终点说道:已经结束了!
吞蛋白介导的快速内吞(Fast Endophilin-Mediated Endocytosis, FEME)之所以快速,是因为它不需要网格蛋白的参与,整个过程没有外衣的装配和解装配,因此速度要快得多。
吞蛋白(Endophilin)是BAR蛋白家族的成员,具有弯曲细胞膜的BAR结构,同时,它还有非常重要的SH3结构域,能够和多种蛋白结合。
吞蛋白有三个功能:
1,通过SH3结构域招募与配体结合的受体;
2,通过BAR结构弯曲细胞膜;
3,通过多个两亲性的螺旋结构帮助动力蛋白切割细胞膜。
吞蛋白介导的快速内吞的第一步,是受体的激活。位于细胞膜表面的受体与配体结合后即会被激活。激活的受体能够通过吞蛋白上的SH3结构域结合,招募吞蛋白,从而起始内吞的过程。
例如,前面提到的β1肾上腺素能受体,它含有脯氨酸富集结构域,在被激活后能够与吞蛋白的SH3结构域结合,从而激活吞蛋白介导的快速内吞,使细胞能够快速回收囊泡。
第二步,吞蛋白通过弯曲细胞膜,将细胞膜向内扯出一个凹陷。
第三步,动力蛋白Dynamin被吞蛋白招募过来,切割与细胞膜连接处的囊泡的脖子。
当然,整个过程中的有多种蛋白和脂质的参与,如板状伪足蛋白(Lamelliphdin)、磷脂酰肌醇等。
总结受体通过结合配体后被激活,被激活的受体又能够与吞蛋白结合,从而引发快速的囊泡回收。这个过程就是吞蛋白介导的快速内吞。
除网格蛋白介导的内吞、吞蛋白介导的快速内吞以外,“吻了就跑”就像是给弹夹一个一个的装子弹。而超快的内吞作用使突触能够以难以置信的速度将所有的空壳迅速填满,然后再排列整齐,使机关枪的弹药永远不会耗尽。后面两种方式回收的速度比吞蛋白介导的快速内吞还要快。
囊泡回收有多种不同的方式,它们速度各异,可以在不同情况下发挥囊泡回收的作用,具体功能在学术界仍有一些争议。
本文内容由快快网络小涵整理编辑!