光合作用是伟大的魔术(光合作用光的作用)
导语:伟大而又精妙的光合作用
叶绿体是植物细胞所特有的能量转换细胞器,其主要功能是进行光合作用。光合作用是自然界最重要的化学反应,是包括人类在内的生物体赖以生存和繁衍的基础,也是能源利用和开发的一个重要方向。全球绿色植物等生物体每年通过光合作用能将太阳能转化为2200亿吨生物能源,相当于全球每年能耗的10倍。如果真的能够大规模仿生利用太阳能,那么困扰人类的食品和能源问题将会有全新的解决方案。
一、叶绿体的形态结构
叶绿体的形状、大小和数量因植物种类不同而有很大差别。同时,叶绿体是一种不稳定的细胞器,能够伴随不同的环境条件,如光照条件等产生不同的适应性变化。高等植物的叶绿体多呈凸透镜状或香蕉形,宽2~4μm,长5~10μm。大多数叶肉细胞含有几十至几百个叶绿体,可占细胞质体积的40%~90%。藻类植物通常只有一个巨大的叶绿体,其形态有带状、螺旋状、星状,依细胞的形态而定。
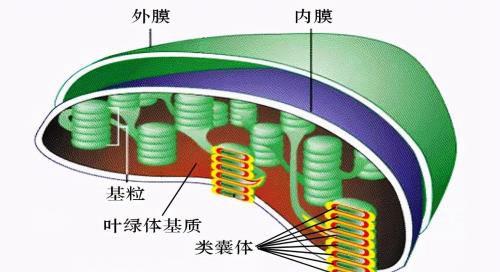
在电子显微镜下可以看到,叶绿体由3部分构成:叶绿体膜、类囊体以及两者之间的空间称为基质。
叶绿体的结构
叶绿体膜由外膜和内膜组成。每层膜厚6~8nm,内、外膜之间为 10~20nm宽的腔隙。与线粒体外膜一样,叶绿体的外膜通透性大,含有孔蛋白,允许相对分子质量高达104的分子通过。叶绿体内膜的通透性相对较差,是细胞质与叶绿体基质间的功能屏障,仅有O2、CO2和H2O分子能自由通过。叶绿体内膜上有很多转运蛋白,它们能够选择性转运进出叶绿体的分子,其运输作用靠浓度梯度驱动。这些转运蛋白的一个重要运输机制是交换转运,如磷酸交换载体,能够将细胞质中的无机磷转运到叶绿体基质,同时将叶绿体基质中产生的磷酸甘油醛释放到细胞质。
叶绿体内部由内膜发展而来的封闭的扁平膜囊,称为类囊体。类囊体囊内空间称为类囊体腔。在叶绿体的某些部位,许多圆饼状的类囊体有序堆叠成垛,称为基粒。一个叶绿体含有40~60个甚至更多的基粒,一个基粒由5~30个类囊体组成,最多可达上百个。组成基粒类囊体的数目,依不同植物或同一植物不同部位的细胞而有很大变化。类囊体堆叠成基粒,是高等植物细胞所特有的膜结构,这种结构大大增加了膜片层的总面积,能更有效地捕获光能,加速光反应。类囊体腔彼此相通,因而一个叶绿体内的全部类囊体实际上是一个完整连续的封闭膜囊,使膜系统的膜囊与基质隔开,这在电化学梯度的建立和ATP的合成中起重要作用。类囊体膜的化学组成与细胞的其他膜成分不同,含极少的磷脂和丰富的具有半乳糖的糖脂,这些脂质中的脂肪酸主要是不饱和的亚麻酸,约占87%。因此,类囊体膜的脂双分子层流动性非常大。脂双分子层的流动性促进光合作用过程中类囊体膜上的光系统Ⅱ、 细胞色素b6/f复合物、光系统I及 ATP合酶复合物在膜上的侧向移动,有利于光合作用的进行。此外,类囊体膜上的蛋白质/脂质比很高,这也与叶绿体行使光合作用有关。
叶绿体内膜与类囊体之间的区室就是叶绿体基质。基质的主要成分是可溶性蛋白质和其他代谢的活跃物质,其中核酮糖-1,5-二磷酸羧化酶/加氧酶是光合作用中一个起重要作用的酶系统,在基质中含量很多,亦是自然界含量最丰富的蛋白质。基质是光合作用固定CO2的场所,基质中还含有环状DNA、核糖体、脂滴、植物铁蛋白和淀粉粒等。
二、光合作用
光合作用是绿色植物利用太阳能将水和二氧化碳转变为有机化合物并放出氧气,将光能转换为化学能的过程,其本质是呼吸作用的逆过程。线粒体中的呼吸作用将氧还原成水,而叶绿体中的光合作用将水氧化产生氧,前一个过程释放能量,后一个过程则需要能量。高等植物的光合作用涉及两个过程:依赖光的光反应和有光无光均可的暗反应。光反应只有在光照下才发生,暗反应(也叫碳反应)是在光反应的产物驱动下进行的。光反应包括水的光解和电子传递及光合磷酸化两个步骤,是在类囊体膜上通过叶绿素等光合色素分子吸收、传递光能,并将光能转换为电能进而转换为活跃的化学能形成ATP和 NADPH,同时也产生O2。暗反应是在叶绿体基质中进行的酶促化学反应,利用光反应产生的ATP和 NADPH,将CO2还原为糖,即将活跃的化学能转换为稳定的化学能,最终储存于有机物中。
(一)光反应
光除去具有波动性外,还有粒子性,也就是说光的行为也像极小的能量“颗粒”一样,这种“颗粒”称为光子。不同波长的光,其光子所带的能量多少不同。每种波长的光,其光子中的能量是固定的。波长越短,光子中的能量越多。例如,紫光的光子中所带的能量约为红光的光子中的两倍。色素吸收光的实质是色素分子中的一个电子得到了光子中的能量。这时这个电子从基态进入激发态,成为一个激发的电子或者说高能电子。所以叶绿素分子吸收光子的一刹那,光子的能量已经变为电子的能量,也就是已经变成了化学能。色素分子的激发态极不稳定,几乎形成后立即变回为基态。叶绿素分子的激发态只维持10-8s。当叶绿素分子从激发态回到基态时,其所吸收的光能便以热的形式向周围发散或转变成荧光。
实验表明,叶绿体中不是所有的叶绿素分子都直接参与光反应将光能转换为化学能。大约300个叶绿素分子组成一个功能单位才能进行光子的吸收,该功能单位称为光系统,它是进行光合作用的最小结构单位。在光系统中只有一对特殊的叶绿素a分子,称为反应中心色素分子,具有将光能转换为化学能的特殊功能,而其余的光合色素称为天线色素分子。它们吸收光能,并将光能迅速而有效地传递到反应中心色素。能量的传递遵守从需能较高的天线色素分子到需能较低的天线色素分子的原则,或者说能量传递的方向是从吸收短波长光波向吸收长波长光波的天线色素分子传递,反应中心的叶绿素分子是吸收光波波长最长的色素,这就使所有天线色素分子吸收的光能必然地、不可逆转地传递给反应中心色素。反应中心色素的最大特点是,在直接吸收光能或接受从天线色素分子传递来的光能而被激发后产生电荷分离和能量转换。
总的来说,光系统是由3个部分组成的:作用中心、天线色素和几种电子载体。这3个部分中都有特定的蛋白质,光系统之所以能在瞬间就抓住激发电子,不使它回到基态去,关键就在于这些蛋白质分子的精巧排列。类囊体膜上有两类光系统,依其发现的先后,分别名为光系统I和光系统Ⅱ。光系统I中的作用中心内的叶绿素a称为P700,因为它与特定的蛋白质结合,其光吸收高峰在700nm,在红光区内。同样,光系统Ⅱ中的叶绿素a称为P680,因为其吸收高峰在680mm,680mm也在红光区,不过稍稍偏向黄橙光。光合膜中有许多个光系统,它们分别组成多个电子传递链。
光系统Ⅱ中的天线色素分子吸收光,并将光能传递给反应中心P680,P680吸收能量后激发出一个电子,传给膜蛋白D1上的原初电子受体脱镁叶绿素(Pheo),然后将电子传给与膜蛋白D2结合的质体醌PQA,再传给与D1蛋白结合的另一个质体醌PQB。PQB在连续接受2个电子后,成为PQB2-,这时PQB2-从基质中摄取2个H+形成还原型PQBH2,PQBH2移到膜的内侧,将2个质子释放到类囊体腔中,而把电子交给细胞色素b6/f,随后又经过位于膜内侧的的质体蓝素(PC),把电子传递给P700。在上述电子传递时,来自水的电子经酪氨酸残基(Tyrz)传递到反应中心P680,水则被氧化分解为O2和H+。
电子传递过程是能量递降的,电子每从一个载体传递到下一个载体时都会丢失部分能量,其中有一些能量就形成了ATP中的能量。这个传递链中的每一步反应都是氧化还原反应。当电子从光系统Ⅱ传递到光系统I的P700时,能量水平已经不高了,不足以还原NADP+,这时被光激发的P700又产生另一个高能电子,其能量水平比光系统Ⅱ中所产生的高能电子还高,于是电子传递继续进行。
光系统I中的天线色素分子吸收光,并将光能传递给反应中心P700,使其中一个叶绿素a分子成为激发态从而释放1个电子,释放的电子传递给原初电子受体——另一个单体叶绿素a分子(A),A又将电子传给A1(可能是叶绿醌 ,即维生素K1),再通过铁硫蛋白将电子传递给铁氧还蛋白(Fd)。Fd最终将电子交给NADP+,NADP+获得2个电子并结合基质中的1个H+后在铁还氧蛋白-NADP+酶的作用下还原成NADPH。这样最终完成了从H2O到NADP+的电子传递。在进行电子传递的同时,H+被泵过类囊体膜,在类囊体膜两侧建立起了质子梯度。
每吸收2个光子(每个光系统各吸收1个),就有1个电子从H2O传递到NADP形成1分子的O2有4个电子从2个H2O传递到2个NADP+,总共必须吸收8个光子其中每个光系统各吸收4个。
在电子传递过程中, 细胞色素b6/f起着质子泵的作用,将H+从叶绿体基质泵到类囊体腔中,水裂解所产生的H+也留在类囊体腔,结果使腔内的H+浓度增加,在类囊体膜两侧建立质子梯度,从而推动H+通过膜中的ATP合酶到达膜外(协助扩散)发生磷酸化作用,使ADP与Pi形成ATP。新合成的ATP立即被释放到基质中。同样,光系统I所形成的 NADPH也在基质中,这样光合作用的光反应产物ATP和NADPH都处于基质中,便于被随后在叶绿体基质中进行的暗反应所利用。
(二)暗反应
高等植物的碳同化共有三条途径:卡尔文(Calvin)循环、C4途径和景天酸代谢途径。其中卡尔文循环是最基本的途径。
卡尔文循环固定CO2的最初产物是甘油酸-3-磷酸(三碳化合物),故也称C3途径。这是20世纪50年代卡尔文等人应用CO2示踪方法所进行的著名实验研究中发现的。由于 Calvin在光合碳同化途径上做出了重大贡献,因此获得了1961年诺贝尔化学奖。C3途径是所有植物进行光合碳同化所共有的基本途径,它包括一系列复杂的反应,但可简化为3个阶段,即羧化(CO2固定)、还原和RuBP再生阶段。
羧化阶段:CO2在被 NADPH还原之前,必须经过羧化阶段固定生成羧酸,然后才被还原。核酮糖-1,5-二磷酸(RuBP)是CO2接受体,在RuBP羧化酶/加氧酶的催化下,1分子RuBP与1分子CO2反应形成1分子不稳定的六碳化合物,并立即分解为2分子甘油酸-3-磷酸。这就是CO2羧化阶段。
还原阶段:甘油酸-3-磷酸在甘油酸-3-磷酸激酶的催化下被ATP磷酸化,形成甘油酸-1,3-二磷酸,然后在甘油醛-3-磷酸脱氢酶催化下被 NADPH还原成储能更多的甘油醛-3-磷酸。这一阶段是耗能反应,光反应中生成的ATP和 NADPH主要是在这一阶段被利用。所以,还原阶段是光反应与暗反应的连接点。一旦CO2被还原成甘油醛-3-磷酸,光合作用的储能过程便完成。甘油醛-3-磷酸等三碳糖可进一步转化,在叶绿体内合成淀粉,也可透出叶绿体,在细胞质中合成蔗糖。
RuBP再生阶段:甘油醛-3-磷酸可异构化,形成磷酸二羟丙酮,这两种磷酸丙糖在醛缩酶催化下合成磷酸己糖。然后在一系列酶的作用下可形成磷酸化的丁糖、戊糖、庚糖,形成的核酮糖-5-磷酸在核酮糖-5-磷酸激酶的催化下发生磷酸化,消耗一个ATP,再形成RuBP,这是第三次耗能的反应。RuBP是卡尔文循环的起始物、CO2的接受体,它必须不断地再生才能维持循环的继续进行。
光合作用的过程
综上所述,C3途径是靠光反应生成的ATP及 NADPH作能源,推动CO2的固定、还原。每循环1次只能固定1个CO2分子,循环6次才能把6个CO2分子同化成1个己糖分子。每固定1个CO2分子,需要3分子ATP和2分子 NADPH。
免责声明:本站部份内容由优秀作者和原创用户编辑投稿,本站仅提供存储服务,不拥有所有权,不承担法律责任。若涉嫌侵权/违法的,请反馈,一经查实立刻删除内容。本文内容由快快网络小奈创作整理编辑!